Here is the description of an organism that is little more than a collection of microtubules
The Slime mold, a single-celled organism that has no brain and no neural network, but it has a cytoskeleton and is filled with cytoplasm both organized by microtubules.
On the role of the plasmodial cytoskeleton in facilitating intelligent behavior in slime mold Physarum polycephalum
Abstract
The plasmodium of slime mold Physarum polycephalum behaves as an amorphous reaction-diffusion computing substrate and is capable of apparently ‘intelligent’ behavior. But how does intelligence emerge in an acellular organism?
Through a range of laboratory experiments, we visualize the plasmodial cytoskeleton—a ubiquitous cellular protein scaffold whose functions are manifold and essential to life—and discuss its putative role as a network for transducing, transmitting and structuring data streams within the plasmodium. Through a range of computer modeling techniques, we demonstrate how emergent behavior, and hence computational intelligence, may occur in cytoskeletal communications networks. Specifically, we model the topology of both the actin and tubulin cytoskeletal networks and discuss how computation may occur therein.
Furthermore, we present bespoke cellular automata and particle swarm models for the computational process within the cytoskeleton and observe the incidence of emergent patterns in both. Our work grants unique insight into the origins of natural intelligence; the results presented here are therefore readily transferable to the fields of natural computation, cell biology and biomedical science. We conclude by discussing how our results may alter our biological, computational and philosophical understanding of intelligence and consciousness.
Introduction
Slime mold Physarum polycephalum’s vegetative life cycle stage, the plasmodium (plural plasmodia, Fig. 1), is a macroscopic, multinucleate, acellular organism which behaves as a living amorphous reaction-diffusion computing substrate.[1]
To delineate, slime mold may be considered as an unconventional computing substrate in which data are represented as transitions in chemical equilibria in an excitable medium. The plasmodium is able to concurrently sense input from a range of stimuli including temperature, light, chemicals, moisture, pH and mechanical force.2-5 P. polycephalum’s innate behavior patterns may be manipulated experimentally to perform useful computational tasks, such as to calculating the shortest route between any number of spatially distributed nutrient sources, or navigating its way out of a maze via chemotaxis; these operations may be interpreted in terms of computational geometry, logic and spatial memory.[6,7]
Figure 1.
Plasmodium of slime mold P. polycephalum growing on an agar-filled Petri dish, feeding on porridge oats. Note the differences in morphology between medial/posterior plasmodial ‘veins’ (black arrow) and the ‘fan-shaped’ anterior margin (white arrow).
We are rapidly approaching the physical limitations of the materials used in the creation of traditional solid state computers, and their manufacture is ecologically-damaging. For these reasons, research into unconventional computing is gathering momentum. Unconventional, or non-classical computation, (UC) utilises the natural properties and processes of physical or living materials to provide useful computational functions. These systems are typically composed of simple and plentiful components, contain redundant parts (i.e. not being dependent on highly complex units), and show resilient or ‘fault tolerant’ behavior. UC is often observed in systems which show ‘emergent behavior’, novel behavior which emerges from the interactions between simple component parts and which—critically—cannot be described in terms of the lower level component interactions. Emergent behavior is found in systems with many simple, local interactions and which display self-organization, i.e. the spontaneous appearance of complexity or order from low-level interactions. Many of the attractive features of UC computing devices (distributed control, redundancy, fault tolerance) are generated by mechanisms of self-organization and emergence, and the study of these properties is useful not only from a computational perspective, but also from a biological viewpoint—since much of the complexity in living systems appears to be built upon these principles.
…
We begin by proposing that the P. polycephalum plasmodium must possess some form of network through which sensory and motor data may travel, as information transmission is a fundamental component of every system capable of computation (partnered with the means for data storage and logical processing of data). Furthermore, without such a network, incoming data—defined here loosely as an environmental stimulus which elicits a response within the organism—would be ‘unstructured’, data structuring here meaning the production of predictable, quantifiable data patterns from unstructured environmental data.
Indeed, in Ref.15, Lungarella and Sporns argue that the dynamical coupling of sensorimotor data streams with the morphology (here meaning both structure and physical properties) of a computational entity’s (biological or artificial) data network induces automatic information structuring, which in turn allows the entity to dynamically react to its environment: this is thought to be the underlying basis of learning and logical ability.[16-19]
Consequently, as the morphology of these data streams define how the entity may sense and interact with its environment, it essentially carries out a proportion of the computation. As such, computational processes are ‘outsourced’ to the morphology automatically, which reduces the workload of the entity’s control unit/s.[15,16,20-22]
This view of natural computing is derived from the precepts of ‘morphological computation’, a concept usually employed in robot design, wherein entity compliance is exploited to create self-stabilizing systems which reduce the need for constant monitoring by the control unit.[16,22]
By amalgamating biological and computer sciences, recent advances in the field have shown promise in modeling natural systems which have not been fully described in purely mathematical (algorithmic) terms, e.g. brain function.[23]
While assigning a tangible structure to the abstract term ‘data stream’ may appear to be a simplification for the ease of description, it is clear that some form of network for optimizing intracellular communication is present within slime mold, as the phenomena we may label as ‘inputs’ (i.e. environmental sensing) display redundancy, rapidity and transduction into different formats. For example, cellular signaling events may be observed to propagate far faster than simple diffusion of signal molecules through the cytoplasm would allow for, and many stimuli are transduced into multiple different formats, e.g., mechanical stimulation of a cell can provoke the generation of biochemical and bioelectrical signaling events.[24]
Slime molds would also appear to have a unique necessity for a data network as a single plasmodium can contain many millions of nuclei. While likening the functions of a cell’s nucleus with those of a processor is a contentious issue (which is not discussed here), it is clear that the nuclear processes which alter the cell/the cell’s behavior (activation/repression of genes, induction of signaling cascades etc.) are direct consequences of the environmental data it receives. How, then, are the activities of millions of nuclei are synchronized to produce coherent behavior? The existence of a plasmodial data network would appear to be an elegant solution to this problem, if such a network were demonstrated to connect nuclei together (hence facilitating internuclear communications) and/or interconnectivity (to allow for signal amplification).
All eukaryotic (and some prokaryotic) organisms possess an intracellular network; a plentiful protein scaffold known as the cytoskeleton, which is composed of tubulin microtubules (MTs), actin microfilaments (MFs) and a range of intermediate filaments (IFs) (Fig. 2). Tubulin and actin are ubiquitous, while IF type differs depending on the function of the cell. All three cytoskeletal protein groups are considered to interlink with each other and with most of the major cellular organelles and receptors, to form one single interconnected network.[25]
The cytoskeleton participates a multitude of cellular functions, many of which are considered to be essential to life; these include mechanical rigidity, motility and substance (organelle and molecular) trafficking.[26]
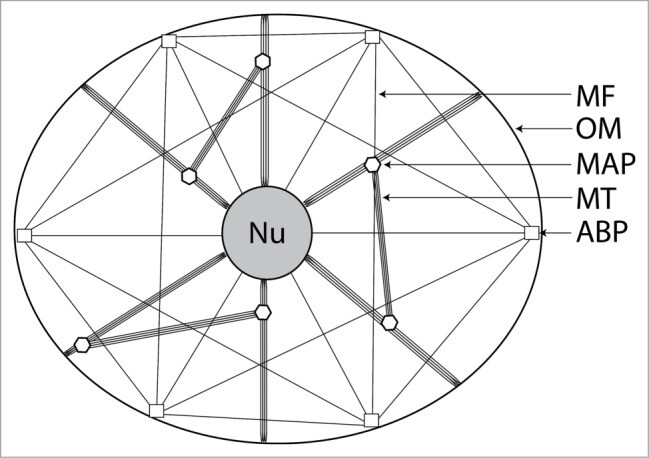
Figure 2.
Simplified schematic representation of the cellular cytoskeleton in a generalized eukaryotic cell, illustrating how several varieties of cytoskeletal protein form an interconnected network which links cellular organelles together. Intermediate filaments and centrosome are not shown. Nu: nucleus; OM: outer membrane; MAP: microtubule-associated protein; MT: microtubule; ABP: actin-binding protein.
It is becoming increasingly evident that the cytoskeleton also participates in a wide range of cellular signaling events. The forms of information it may transmit include electrical potential, mechanical force, quantum events, propagating waves of protein conformational changes, facilitated chemical transport and biochemical signal transduction cascades.[24,26-32]
The cytoskeleton therefore represents an attractive model for describing data transmission within cells, as all of these signaling events may be interpreted as data being fed into a cellular computer. We are by no means the first to suggest this, but such a topic has not been previously described in slime mold models.
This study was undertaken to explore the putative role of the slime mold cytoskeleton in facilitating intelligent behavior by acting as an intraplasmodial data network. This was achieved by visualizing the plasmodial cytoskeleton with confocal microscopy and basing a range of computer modeling techniques on the structural observations made.
Results
Visualizing and formalising the plasmodial cytoskeleton
The actin and tubulin components of the P. polycephalum cytoskeleton were visualized with confocal microscopy, see Figures 3 and 4 (see Methodology section for details of sample preparation). Samples were taken from the 2 distinct anatomical locations of the plasmodium, namely the tubular plasmodial ‘veins’ that form the posterior areas of the plasmodium and the ‘fan-shaped’ advancing anterior margin formed from converging pseudopodia (see Fig. 1). The actin and tubulin components of the plasmodial cytoskeleton appear to be extremely complex, highly interconnected networks – especially so when compared to animal cell counterparts. The actin MF network is extremely abundant in the advancing pseudopodia (likely to result from the propulsion of motile machinery to the pseudopodial growth cone to facilitate movement), although somewhat less so in plasmodial veins (data not shown); the tubulin network would appear to be profuse in both anatomical locations, but less so than actin in the anterior portions of the organism. Both cytoskeletal proteins appear to articulate onto each nucleus at several points and are also present at the external membrane, indicating associations with surface receptors (as has been demonstrated in animal cells).26
more…
What does this suggest about microtubules???
Moreover, I can see no more persuasive argument that microtubules are the prime constituent of “intelligent networks” than that even single-celled brainless and neuronless organisms have inherent “communication” skills that may be very much like “quorum sensing” in bacteria, plants and eventually acquire emergent self-aware conscious intelligence in more evolved complex animals.
An organism does not need to be conscious to be sentient and responsive to survival demands.
Consciousness is the evolutionary finish of increasingly complex sensory response networks and emerges at every level of natural selection of survival strategies.



 James E. Trosko
James E. Trosko

{kind=link}